Thierry Douki & Jean-Luc Ravanat UV induced DNA damage plays a key role in the initiation phase of skin cancer and is involved in other
photobiological responses. When DNA damage is left unrepaired or when damaged cells are not eliminated by apoptosis, DNA lesions express their mutagenic properties. This process may lead to the activation of proto-oncogene or the inactivation of tumor suppression genes. The chemical nature and the amount of DNA damage strongly depend on the wavelength of the incident photons. The most energetic part of the solar spectrum at the Earth’s surface (UVB, 280 320 nm) leads to the formation of
DNA photoproducts by cycloaddition reactions between adjacent pyrimidine bases. Cyclobutane pyrimidine dimers (CPDs) involve cycloaddition of the two C5-C6 double bond of two pyrimidine rings while pyrimidine (6-4) pyrimidone photoproducts (64PP) result from the addition of the C5-C6 double bond of 5’-end base to the carbonyl or amino group of the 3’-end base. Upon exposure to large doses of UVB or to UVA, 64PP undergo photoconversion onto their Dewar valence isomers. This photoreactivity of DNA is similar in all cell types (mammals, plants bacteria, etc.). One exception are bacterial spores where DNA is surrounded by specific proteins and is in a dehydrated state. This results in a drastic change in its conformation and the only photoproduct formed is a “spore photoproduct” that corresponds to the formation of a methylene bridge between adjacent thymines. CIBEST is actively involved in understanding the parameters that affect the distribution of these pyrimidine dimers. Investigations rely on the
HPLC-mass spectrometry detection of the photoproducts. Its versatility allows to quantify all possible dimers at each of the four bipyrimidine sites in a sample and in in single analysis. Moreover, the high sensitivity allows us to work at low dose both in test tube experiments to study basic photochemical events and in cells or skin to mimic actual exposures. The HPLC MS/MS detection of photoproducts is also a valuable tool in the study of
DNA repair. In these works, we use various types of
UV lamps and a solar simulator.
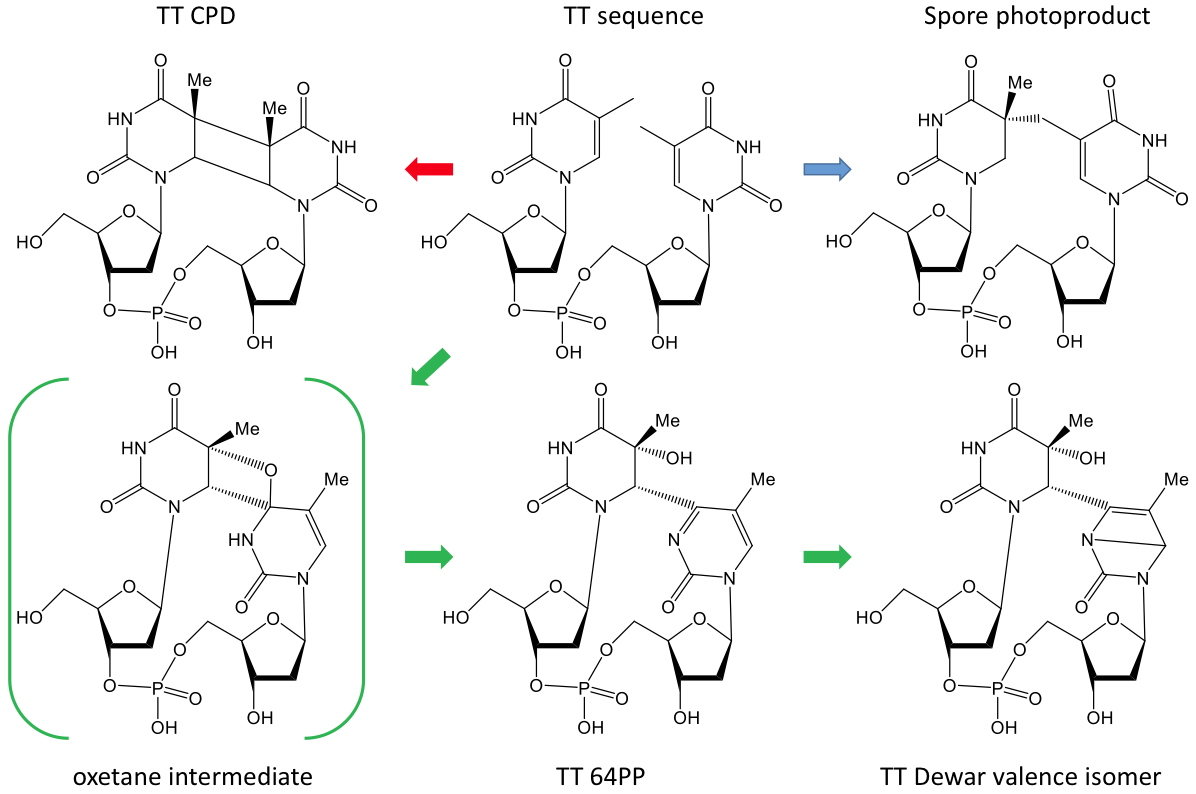
Chemical structure of pyrimidine dimers.
The figure show thymine-thymine derivatives. With the exception of the spore photoproducts, similar dimers are produced, although in different yields, at thymine-cytosine, cytosine-thymine and cytosine-cytosine sequences.
Long neglected or considered mainly for its involvement in skin aging, UVA is now recognized as being capable of inducing mutations and tumors. Although its effectiveness is less than that of UVB, UVA rays are at least 20 times more abundant. The major effect of UVA in skin cells is the production of
oxidizing chemical species, such as the highly mediatized “free radicals” or the most discreet but equally effective “singlet oxygen“, an activated form of molecular oxygen. These compounds are produced by
photosensitization processes that are triggered when certain cellular components absorb UVA and transfer the absorbed energy to oxygen or remove electrons from nearby molecules. The reactive oxygen species thus generated can then damage the cellular macromolecules: lipids, proteins and DNA. The nature of DNA oxidation products is very varied and depends on the reactive oxygen species involved. Unraveling the chemical reactions leading to oxidatively generated DNA lesions is a major line of research at CIBEST. The group has provided large amounts of information on the reactivity of DNA with singlet oxygen. It is now well established that only guanine is affected and that only 8-oxoGua is generated. Other DNA lesions can be produced but they are biologically irrelevant over-oxidation products. Production of singlet oxygen is not the only photosensitized oxidation mechanism. Type I processes involving one-electron abstraction can also be triggered. They are yet a minor component of the oxidative stress induced to DNA upon exposure of cells to UVA. Such processes remain of interest because they are involved in the phototoxic effects of some drugs. Another major ROS involved in the effects of UVA is the hydroxyl radical •OH, which is well known in the effect of
ionizing radiations and
metals. In contrast to singlet oxygen, •OH reacts with the four DNA bases as well as 2’-deoxyribose units. Consequently, a wide variety of oxidation products are generated together with single strand breaks. The distribution of oxidatively generated DNA lesion in UVA-irradiated cells showed that singlet oxygen accounts for approximately 80% of the damage. The reality of the effects of UVA is more complex than the mere induction of oxidative stress as it is often highlighted. Indeed, it has been shown in recent years that UVA can also induce pyrimidine dimers in DNA. Although the yield is much lower than with UVB, dimers are still more frequent than oxidation products in the DNA of cells exposed to UVA. Finally, it has been reported that in human cells, UVA, like UVB, induces mutations mainly at sites containing two pyrimidines. Although UVA is less harmful than UVB, its contribution to the genotoxic properties of sunlight cannot be neglected. In particular, these data must be taken into account when considering a reduction in the use of artificial tanning lamps, which are very rich in UVA. Their use can be considered as a voluntary exposure to a mutagenic and carcinogenic agent with no health benefit.
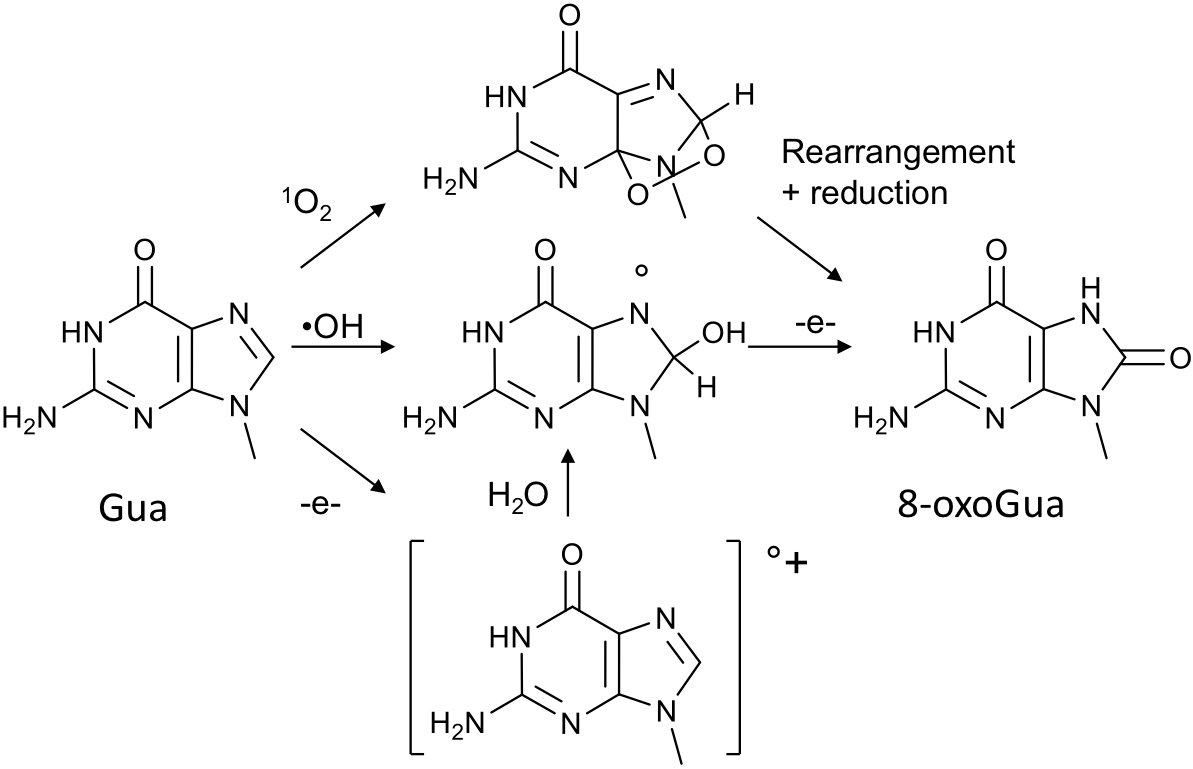
The different mechanisms leading to the formation of 8-oxoGua.